Inhibitory Effect of an Ethanol Extract Mixture of Vitis amurensis, Aralia cordata, and Glycyrrhizae radix on Amyloid β Protein (25-35)-Induced Neurotoxicity
© The Korean Society of Medicinal Crop Science
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License ( http://creativecommons.org/licenses/by-nc/3.0 ) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
The present study investigated an ethanol extract (SSB) of a mixture of three medicinal plants of Vitis amurensis, Aralia cordata, and Glycyrrhizae radix for possible neuroprotective effects on neurotoxicity induced by Amyloid β protein (Aβ) (25-35) in cultured rat cortical neurons and antidementia activity in mice. Exposure of cultured cortical neurons to 15 μM Aβ (25-35) for 36 h induced neuronal apoptotic death. At 1-30µg/ml, SSB inhibited neuronal death, elevation of intracellular calcium concentration ([Ca2+]i), and generation of reactive oxygen species (ROS) induced by Aβ (25-35) in cultured cortical neurons. Memory impairment and increase of acetylcholinesterase activity induced by intracerebroventricular injection of mice with 16 nmol Aβ (25-35) was inhibited by chronic treatment with SSB (25, 50 and 100 mg/㎏, p.o., for 8 days). From these results, it is suggested that antidementia effect of SSB is due to its neuroprotective effect against Aβ (25-35)-induced neurotoxicity and that SSB may have a therapeutic role in preventing the progression of Alzheimer’s disease.
Keywords:
Vitis amurensis, Aralia cordata, Glycyrrhizae radix, Amyloid β Protein, Cultured Neurons, Memory ImpairmentINTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disorder, and the most common type of dementia. It is characterized by pronounced loss of neurons, extracellular senile plaque, whose principle component is β-amyloid (Aβ), a 39-43 amino acid peptide derived from the cleavage of the amyloid precursor protein by β and γ-secretase (Ivins et al., 1999). Aβ can directly induce neuronal cell death (Ueda et al., 1997) and cause neurons vulnerable to excitotoxicity (Koh et al., 1990) and oxidative insults (Chen et al., 2007). It is suggested that the mechanisms underlying Aβ neurotoxicity are complex but may involve N-methyl-D-aspartate (NMDA) receptor, a glutamate receptor subtype, modulation induced by glutamate release, sustained elevations of intracellular Ca2+ concentration ([Ca2+]i), and oxidative stresses resulting from reactive oxygen species (ROS) generation (Ekinci et al., 2000; Gray and Patel, 1995; Ueda et al., 1997). Aβ has been reported to produce hydrogen peroxide, superoxide, and proinflammatory cytokines in neurons and glial cells (Gitter et al., 1995). Therefore, antioxidants such as α-tocopherol and antiinflammatory agents such as indomethacin reportedly slow the progression of AD (Gasparini et al., 2004; Sano et al., 1997).
Vitis, known as a kind of grapes, has various pharmacological effects including improvement against fatty liver, hypertension, cardiac diseases and arthritis (Wang et al., 2004). Vitis contains large amount of polyphenolic compounds such as resveratrol possessing potent antioxidant properties (Rice-Evans et al., 1995) and anti-inflammatory properties (Wang et al., 2004). Aralia belongs to the families Araliaceae and has long been recognized as therapeutic herbs for antinociceptive, antidiabetic, antioxidant and anti-inflammatory activities in China, Japan and Korea (Bae, 2004). Diterpenes isolated from the root of this plant were known as active constituents for analgesic and anti-inflammatory agents (Han et al., 1985). Recent studies have indicated that extracts from the aerial parts of A. cordata inhibit cyclo-oxygenase (COX)-1, COX-2, and COX-2-dependent prostaglandin E2 (PGE2) generation, as well as hyperalgesia during peripheral inflammation (Dang et al., 2005; Lee et al., 2006; Park et al., 2005). Glycyrrhizae radix, the root of Glycyrrhizae uralensis (Lycophodiaceae), has been widely used in Chinese medicine and food. Current European and Chinese texts list Glycyrrhizae radix as an expectorant in the treatment of bronchitis, catarrh, and coughs, as an anti-inflammatory agent in gastric ulcers, arthritis, and rheumatism, and as an adrenocorticotropic agent in adrenocorticoid insufficiency (Huang, 1999). Moreover, Glycyrrhizae radix has been reported to have antioxidant effects (Tang et al., 2004).
There has been much effort to develop beneficial agents from medicinal plants with free radical scavenging and anti-inflammatory properties to achieve neuroprotection in our laboratory (Jung et al., 2012; Kim et al., 2011). In our previous studies, aerial parts, leaves and stems, of Vitis amurensis and Aralia cordata were demonstrated to show neuroprotective effect against Aβ (25-35)-induced neurotoxicity in in vivo and in vitro (Cho et al., 2009; Jeong et al., 2010). Glycyrrhizae radix also showed neuroprotection in cultured neurons, but not in in vivo (Lee et al., 2012). Therefore, we hypothetized that mixture of three plants might show more effective and synergistic neuroprotective effect with the reduction of possible individual toxicity due to large quantity.
The present study investigated the neuroprotective property of an ethanol extract of mixture of Vitis amurensis, Aralia cordata, and Glycyrrhizae radix, which was named as SSB, against Aβ (25-35)-induced memory deficits in mice and neuronal damage in cultured rat cortical neurons.
MATERIALS AND METHODS
1. Plant Materials and Extraction
The leaves and stems of V. amurensis and the aerial parts of A. cordata were collected at Keryong Mountain in Daejeon, Korea, from May to July and Glycyrrhizae radix (Korean) was purchased from Daegu Oriental Pharm Co. at Daegu, Korea. These plants were identified by Dr. KiHwan Bae, Chungnam National University. Each 200 g of the three plants was mixed, extracted with ethanol (3 L × 24 h × 3) at room temperature, filtered, and concentrated under reduced pressure using a rotary evaporator to yield an ethanol extract (SSB, 64 g) (Choung et al., 2013), which was then stored at −20°C until required.
2. Experimental Animals
Pregnant Sprague-Dawley (SD) rats for primary neuronal culture and male ICR mice for the passive avoidance test were supplied by Daehan BioLink Co., Ltd. (Chungbuk, Korea) and housed in an environmentally controlled room at 22 ± 2°C, with a relative humidity of 55 ± 5%, a 12-h light/dark cycle, and food and water ad libitum. The procedures involving experimental animals complied with the animal care guide lines of the National Institutes of Health and the animal ethics committee of Chungbuk National University.
3. Induction of Neurotoxicity in Primary Cultures of Rat Cerebral Cortical Neurons
Primary cortical neuron cultures were prepared using embryonic day 15 to 16 SD rat fetuses, as previously described (Ban et al., 2006). Neurotoxicity experiments were performed on neurons after 3-4 days in culture. Cultured neurons were treated with 15 μM Aβ (25-35) (Bachem, Bubendorf, Switzerland) in serum-free DMEM (Sigma, St. Louis, MO, USA) at 37°C for 36 h (unless otherwise indicated) to produce neurotoxicity. An Aβ (25- 35) stock solution of 2 mM was prepared in sterile distilled water, stored at −20°C, and incubated for more than 2 days at 37°C to aggregate before use. SSB was dissolved in dimethyl-sulfoxide (DMSO) at concentrations of 50mg/㎖ and further diluted in experimental buffers. The final concentration of DMSO was less than 0.1%, which did not affect cell viability. For each experiment, SSB was applied 20 min prior to treatment with 15 μM Aβ (25-35). It was also present in the medium during Aβ (25-35) incubation.
4. Measurements of Aβ (25-35)-induced Neuronal Death and Intracellular Biochemical Changes
A 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT, Sigma, St. Louis, MO, USA) assay and Hoechst 33342 (Molecular Probes, Eugene, OR, USA) staining were performed to measure neuronal death 36 h after exposure of cultured neurons to 15 μM Aβ (25-35), as previously described (Ban et al., 2006). Changes in [Ca2+]i were measured with Fluo-4 AM (Molecular Probes), a Ca2+-sensitive fluorescent dye, using a laser scanning confocal microscope (LSM 510, Carl Zeiss, Oberkochen, Germany) with 488-nm excitation argon laser and 515-nm longpass emission filters. The microfluorescence of 2’,7’-dichlorofluorescein, the fluorescent product of 2’,7’-dichlorofluorescein diacetate (H2DCF-DA; Molecular Probes), and a laser scanning confocal microscope (MRC 1024ES, Biorad, Maylands, UK) with 488-nm excitation and 510-nm emission filters were used to monitor the generation of ROS in neurons treated with 15 μM Aβ (25-35) for 24 h.
5. Measurement of Memory Impairment and Brain Cholinesterase Activity
Intracerebroventricular (i.c.v.) injection of the aggregated Aβ (25-35) (16 nmol) was performed to induce memory impairment in mice, as previously described (Kim et al., 2009). SSB (25, 50 and 100mg/㎏) suspended in distilled water was orally administered to 5-week-old ICR mice 30 min before the injection of Aβ (25-35) and further administered daily for 7 days. Passive avoidance apparatus (Gemini Avoidance System, SanDiego, CA, USA) was used to measure memory acquisition according to the method previously described (Kim et al., 2009). Mice were trained on step-through passive avoidance task 30 min after administration of SSB on day 7 of i.c.v. injection of Aβ (25-35). Retention trial was given 24 h after the acquisition trial. Cholinesterase activity of mice whole brain isolated immediately after the retention trial of passive avoidance test was determined using the spectrophotometric method (Ellman et al., 1961).
6. Statistical Analysis
Data were expressed as mean ± SEM and statistical significance was assessed by one-way analysis of variance (ANOVA) and Tukey’s tests. P values of < 0.05 was considered significant.
RESULTS
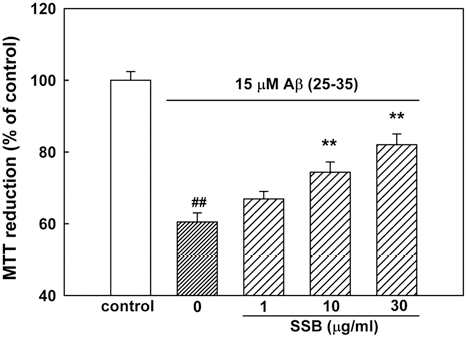
1. SSB Inhibited Aβ (25-35)-Induced Neuronal Cell Death
Cultured cortical neurons exposed to 15 μM Aβ (25-35) for 36 h showed 60.5 ± 2.5% absorbance of that of the untreated controls in the MTT assay (Fig. 1), indicating that Aβ (25-35) caused neuronal cell death. Pretreatment of cortical neurons with 10 and 30µg/ml SSB reduced the neuronal death induced by Aβ (25-35) (absorbance, 74.4 ± 2.8% and 82.0 ± 3.0% of control, respectively; Fig. 1).
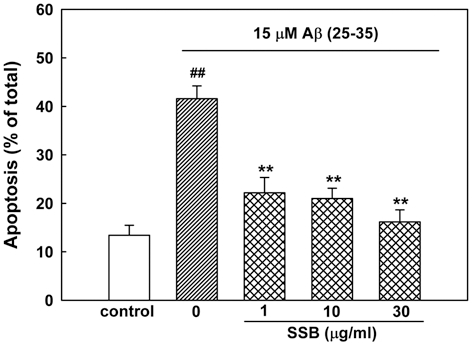
An additional experiment was performed with Hoechst 33342 staining to detect condensed or fragmented DNA, which is indicative of Aβ (25-35)-induced neuronal apoptotic death. Treatment of neurons with 15 μM Aβ (25-35) produced apoptosis of 41.6 ± 2.7% of the total population of cultured cortical neurons, as compared with 13.4 ± 2.1% of apoptotic neurons in control cultures. The addition of SSB (1, 10 and 30µg/ml) significantly decreased the Aβ (25-35)-induced apoptotic cell death, showing 22.2 ± 3.2, 21.0 ± 2.1 and 16.2 ± 2.5%, respectively (Fig. 2).
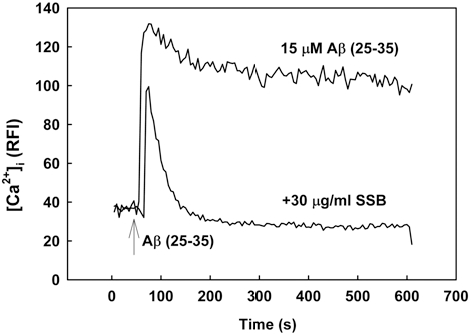
2. SSB Inhibited Aβ (25-35)-Induced [Ca2+]i Elevation
Increase in [Ca2+]i has been associated with Aβ-induced cell death. In cultured neurons, [Ca2+]i rapidly increased in response to treatment with 15 μM Aβ (25-35) and then showed a slow and gradual decrease over 10 min (Fig. 3). In contrast, pretreatment with SSB (30µg/ml) showed inhibition of the increase of [Ca2+]i induced by 15 μM Aβ (25-35).
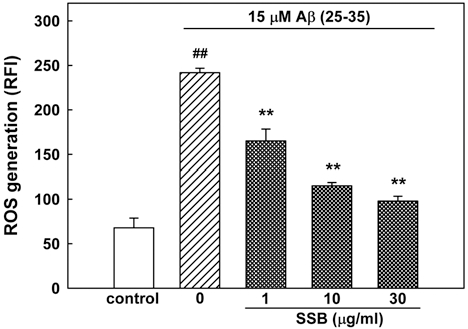
3. SSB inhibited Aβ (25-35)-Induced ROS Generation
To explain the involvement of oxidative stress in Aβ neurotoxicity, the accumulation of ROS after the exposure of the neurons to Aβ (25-35) for 24 h was measured. In H2DCF-DA-loaded cortical neurons, 15 μM Aβ (25-35) increased the fluorescence intensity, indicating that ROS were generated. In neurons treated with 15 μM Aβ (25- 35), the relative fluorescence increased approximately 3.5- fold to 241.8 ± 5.0 compared with the value in control neurons (67.8 ± 10.9, Fig. 4). The Aβ (25-35)-induced increase in ROS generation was significantly inhibited by SSB (1, 10 and 30µg/ml).
4. SSB Inhibited Aβ (25-35)-Induced Memory Impairment
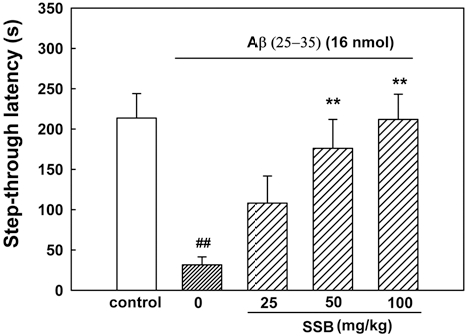
In the initial acquisition trial of the passive avoidance task, the step-through latency did not differ among the 5 groups (control, 16 nmol Aβ (25-35), Aβ (25- 35) + 25mg/㎏ SSB, Aβ (25-35) + 50mg/㎏ SSB, and Aβ (25-35) + 100mg/㎏ SSB; data not shown). The stepthrough latency of the Aβ (25-35)-treated group in the retention trial significantly decreased to 31.6 ± 9.9 s, compared with 213.6 ± 30.3 s in the control group, indicating that Aβ (25-35) impaired memory in mice. Chronically administered SSB markedly protected against the memory impairment produced by Aβ (25-35). The step-through latency in groups administered SSB was 108.1 ± 33.6, 176.0 ± 35.9 and 212.0 ± 31.2 s at doses of 25, 50 and 100mg/㎏, respectively (Fig. 5).
Cholinesterase activity in brain exposed to 16 nmol Aβ (25-35) was significantly increased. In the group consuming 50mg/㎏ and 100mg/㎏ SSB, the cholinesterase activity significantly decreased compared with the Aβ (25- 35) group (Table 1).
DISCUSSION
Aβ (25-35), which is the core toxic fragment of full length Aβ (1-40), forms a β-sheet structure and induces neuronal cell death, neuritic atrophy, synaptic loss, and memory impairment (Maurice et al., 1996; Pike et al., 1995; Tohda et al., 2004). It has been reported that Aβ (25-35) causes neuronal cell death as shown in the present study, and it was blocked by MK-801, an NMDA receptor antagonist; verapamil, a L-type Ca2+ channel blocker; and and NG-nitro-L-arginine methyl ester (LNAME), a nitric oxide synthase inhibitor, in cultured neurons (Cho et al., 2009; Jeong et al., 2010). These results suggest that NMDA glutamate receptor activation, Ca2+ influx through L-type voltage dependent Ca2+ channel (L-VDCC) and ROS generation are implicated in Aβ- induced neuronal apoptotic death. The present study provides evidence that Aβ (25-35)-induced injury to rat cortical neurons was prevented by SSB, an ethanol extract of a mixture of Vitis amurensis, Aralia cordata, and Glycyrrhizae radix. SSB was able to reduce the [Ca2+]i increase, ROS generation, and apoptotic neuronal death in Aβ (25-35)-induced neurotoxicity. Additionally, SSB prevented memory loss induced by i.c.v. injection of Aβ (25-35) in mice.
Ca2+ influx via L-VDCC is considered as a primary insult after Aβ treatment in cultured neurons, because blockade of this channel or Ca2+ chelation prevents other consequences (Ueda et al., 1997). In the present study, SSB inhibited a significant increase of [Ca2+]i after the treatment with Aβ (25-35). Many reports have demonstrated the role of ROS formation in Aβ-induced neurotoxicity (Miranda et al., 2000; Morais Cardoso et al., 2002). It has been reported that vitamin-E, an antioxidant, blocked the Aβ-induced generation of ROS, but not Ca2+ influx, and reduction of extracellular Ca2+ inhibited the Aβ-induced increase in [Ca2+]i as well as generation of ROS, indicating that ROS generation is a consequence of Ca2+ accumulation (Ekinci et al., 2000). SSB also decreased the Aβ (25-35)-induced increase of ROS generation. These results indicate that SSB might prevent Aβ (25-35)-induced Ca2+ entry through VDCC- and / or NMDA-receptor-coupled channels to inhibit ROS generation and then neuronal death, although the mechanism by which SSB blocks the channels is not clear. Vitis amurensis, Aralia cordata, and Glycyrrhizae radix, the constituents of SSB, have been reported to possess antioxidant principles, resveratrol, oleanolic acid, and isoliquirtigenin (Du and Ko, 2006; Ha et al., 2009a, b; Kumar et al., 2007), respectively, suggesting that inhibition of Aβ (25-35)-induced neuronal death by SSB might be due to their ROS scavenging activity. Further study should be conducted to elucidate the precise mechanism.
Many studies have demonstrated that Aβ is accompanied by multiple events culminating in apoptosis in in vitro neuronal experiments (Ekinci et al., 2000; Yan et al., 1999 ). In the present work, cultured cortical neurons exposed to Aβ (25-35) for 36 h showed increased chromatin condensation, a typical feature of apoptotic cell death, which was reduced by SSB. Activation of caspases after increased [Ca2+]i signaling and ROS generation, or inflammatory responses in Aβ-stimulated neurons have been proposed to play pivotal roles in apoptosis. We have already demonstrated that an increase of caspase-3 activity in Aβ (25-35)-treated cultured cortical neurons is correlated with the increase of [Ca2+]i, ROS generation and neural apoptotic death (Ban et al., 2006; Lee et al., 2005). In the previous studies, Vitis amurensis, Aralia cordata, and Glycyrrhizae radix inhibited caspase activity to reduce Aβ (25-35)-induced neuronal apoptosis (Cho et al., 2009; Jeong et al., 2010; Lee et al., 2012). Therefore, SSB might work in the same way.
I.c.v. administration of Aβ (25-35) into mouse brain induces memory impairment in different behavioral paradigms, including spontaneous alternation, the water maze, and passive avoidance (Maurice et al., 1996; Um et al., 2006). Aβ (25-35) preferentially impairs spatial and non-spatial short-term memory, and these effects remain evident up to 6 months after even a single i.c.v. injection of the peptide (Stepanichev et al., 2003). In the present work, memory impairment in the passive avoidance test was also confirmed in mice 7 days after the i.c.v. injection of Aβ (25-35). Chronic treatment with SSB effectively protected the mice against Aβ (25-35)-induced memory deficit. This result was essentially in agreement with its protective effect on Aβ (25-35)-induced neurotoxicity in vitro. Aβ accumulation associated with cognitive impairment in AD is accompanied by an increase in cholinesterase activity (Atack et al., 1983). Equivalently, the increase of cholinesterase activity by Aβ (25-35) in the brain was inhibited by SSB in the present study.
Many reports have suggested that oxidative stress is responsible for the onset of the cognitive dysfunction as well as the progression of AD (Butterfield et al., 2001; Kontush, 2001). Elevated levels of Aβ induce oxidative stress, increasing the appearance of ROS such as superoxide and NO and subsequently producing ONOO- by a rapid interaction, could mediate the damage seen in AD (Kontush, 2001; Smith et al., 1997). A scavenger of ONOO− protects against Aβ (25-35)-induced memory impairment (Alkam et al., 2007), and anti-oxidants such as α-tocopherol protect against cytotoxicity in vitro as well as against learning and memory deficits induced by Aβ (Sano et al., 1997). In the present study, 15 μM Aβ (25- 35) significantly increased the ROS level in cultured neurons, and this was inhibited by SSB. In addition, the constituents of SSB contain antioxidant components (Du and Ko, 2006; Ha et al., 2009a,b; Kumar et al., 2007). Therefore, it is possible that the favorable effect of SSB on Aβ (25-35)-induced cognitive deficits can be attributed to the inhibition of ROS generation.
In the previous studies (Cho et al., 2009; Jeong et al., 2010; Lee et al., 2012), Vitis amurensis, Aralia cordata, and Glycyrrhizae radix were demonstrated to show neuroprotective effect against Aβ (25-35)-induced neurotoxicity in cultured neurons. Furthermore, Vitis amurensis (Jeong et al., 2010) and Aralia cordata (Cho et al., 2009) but not Glycyrrhizae radix, at the dosage of 50 and 100mg/㎏ revealed protection against Aβ (25-35)-induced memory impairment in mice. Mixture of three plants, SSB, showed a maximal inhibition of Aβ (25-35)-induced memory impairment by a dosage of 100mg/㎏ in the present study. These results indicate that three medicinal plants acted synergistically to produce neuroprotection, since maximal effect of SSB was shown at much lower concentration than the sum of maximal effect concentration of each plant.
In conclusion, it is evident that SSB could provide a marked protection against Aβ (25-35)-induced neuronal cell damage in vitro and Aâ (25-35)-induced memory deficit in vivo. These may explain the inhibitory action of SSB on the progression of AD.
ACKNOWLEDGEMENTS
This work was supported by the research grant of the Chungbuk National University in 2012.
REFERENCES
- Alkam, T, Nitta, A, Mizoguchi, H, Itoh, A, Nabeshima, T, A natural scavenger of peroxynitrites, rosmarinic acid protects against impairment of memory induced by Abeta(25-35), Behavioural Brain Research, (2007), 180, p139-145.
-
Atack, JR, Perry, EK, Bonham, JR, Perry, RH, Tomlinson, BE, Blessed, G, Fairbairn, A, Molecular forms of acetylcholinesterase in senile dementia of Alzheimer type Selective loss of the intermediate(10S) form, Neuroscience Letters, (1983), 40, p199-204.
[https://doi.org/10.1016/0304-3940(83)90302-6]

- Bae, K, The medicinal plants of Korea, (eoul), Seoul, Korea, Kyo-Hak Publishing Co, p362-363.
- Ban, JY, Jeon, SY, Bae, K, Song, KS, Seong, YH, Catechin and epicatechin from Smilacis chinae rhizome protect cultured rat cortical neurons against amyloid beta protein(25- 35)-induced neurotoxicity through inhibition of cytosolic calcium elevation, Life Sciences, (2006), 79, p2251-2259.
- Butterfield, DA, Drake, J, Pocernich, C, Castegna, A, Evidence of oxidative damage in Alzheimer's disease brain: Central role for amyloid beta-peptide, Trends in Molecular Medicine, (2001), 7, p548-554.
-
Chen, CY, Jang, JH, Park, MH, Hwang, SJ, Surh, YJ, Park, OJ, Attenuation of abeta-induced apoptosis of plant extract(Saengshik) mediated by the inhibition of mitochondrial dysfunction and antioxidative effect, Annals of the New York Academy of Sciences, (2007), 1095, p399-411.
[https://doi.org/10.1196/annals.1397.043]
- Cho, SO, Ban, JY, Kim, JY, Jeong, HY, Lee, IS, Song, KS, Bae, K, Seong, YH, Aralia cordataprotects against amyloid beta protein(25-35)-induced neurotoxicity in cultured neurons and has antidementia activities in mice, Journal of Pharmacological Sciences, (2009), 111, p22-32.
-
Choung, MG, Hwang, YS, Kim, GP, Ahn, KG, Shim, HS, Hong, SB, Choi, JH, Yu, CY, Chung, IM, Kim, SH, Lim, JD, Antimelanogenic effect and whitening of anthocyanin rich fraction from seeds ofLiriope platyphylla, Korean Journal of Medicinal Crop Sciences, (2013), 21, p361-371.
[https://doi.org/10.7783/kjmcs.2013.21.5.361]
-
Dang, NH, Zhang, X, Zheng, M, Son, KH, Chang, HW, Kim, HP, Bae, K, Kang, SS, Inhibitory constituents against cyclooxygenases fromAralia cordataThunb, Archives of Pharmacal Research, (2005), 28, p28-33.
[https://doi.org/10.1007/bf02975131]
- Du, Y, Ko, KM, Oleanolic acid protects against myocardial ischemia-reperfusion injury by enhancingtochondrial antioxidant mechanism mediated by glutathione and alpha-tocopherol in rats, Planta Medica, (2006), 72, p222-227.
- Ekinci, FJ, Linsley, MD, Shea, TB, Beta-amyloid-induced calcium influx induces apoptosis in culture by oxidative stress rather than tau phosphorylation, Brain Research, (2000), 76, p389-395.
- Gasparini, L, Ongini, E, Wenk, G, Non-steroidalantiinflammatory drugs(NSAIDs) in Alzheimer's disease: Old and new mechanisms of action, Journal of Neurochemistry, (2004), 91, p521-536.
-
Gitter, BD, Cox, LM, Rydel, RE, May, PC, Amyloid beta peptide potentiates cytokine secretion by interleukin-1 beta-activated human astrocytoma cells, (1995), Proceedings of the National Academy of Sciences of the United States of America, 92, p10738-10741.
[https://doi.org/10.1073/pnas.92.23.10738]
- Gray, CW, Patel, AJ, Neurodegeneration mediated by glutamate and beta-amyloid peptide: A comparison and possible interaction, Brain Research, (1995), 691, p169-179.
- Ha, DT, Chen, QC, Hung, TM, Youn, UJ, Ngoc, TM, Thuong, PT, Kim, HJ, Seong, YH, Min, BS, Bae, K, stillbenes and oligostilbenes from leaf and stem ofVitis amurensisand their cytotoxic activity, Archives of Pharmacal Research, (2009a), 32, p177-183.
-
Ha, DT, Kim, H, Thuong, PT, Ngoc, TM, Lee, I, Hung, ND, Bae, K, Antioxidant and lipoxygenase inhibitory activity of oligostilbenes from the leaf and stem ofVitis amurensis, Journal of Ethnopharmacology, (2009b), 125, p304-309.
[https://doi.org/10.1016/j.jep.2009.06.019]
-
Han, B, Woo, E, Park, M, Han, Y, Studies on the antiinflammatory activity ofAralia continentalis(III), Archives of Pharmacal Research, (1985), 8, p59-65.
[https://doi.org/10.1007/bf02912223]
- Huang, KC, The pharmacology of Chinese herbs, (1999), Boca Raton, Florida, USA, CRC press LLC, p364-368.
- Ivins, KJ, Ivins, JK, Sharp, JP, Cotman, CW, Multiple pathways of apoptosis in PC12 cells. CrmA inhibits apoptosis induced by beta-amyloid, The Journal of Biological Chemistry, (1999), 274, p2107-2112.
- Jeong, HY, Kim, JY, Lee, HK, Hado, T, Song, KS, Bae, K, Seong, YH, Leaf and stem ofVitis amurensisand its active components protect against amyloid beta protein(25-35)-induced neurotoxicity, Archives of Pharmacal Research, (2010), 33, p1655-1664.
-
Jung, MH, Song, KS, Seong, YH, Inhibitory effect of Chaenomeles sinensis fruit on amyloid β protein(25-35)-induced neurotoxicity in cultured neurons and memory impairment in mice, Korean Journal of Medicinal Crop Sciences, (2012), 20, p8-15.
[https://doi.org/10.7783/kjmcs.2012.20.1.008]
- Kim, JY, Kweon, K, Lee, HK, Kim, SH, Yoo, JD, Bae, KH, Seong, YH, Protective effect of an ethanol extract mixture ofCurcuma longaeradixPhellinus linteus, andScutellariaeradix on oxidative neuronal damage, Korean Journal of Medicinal Crop Sciences, (2011), 19, p31-37.
- Koh, JY, Yang, LL, Cotman, CW, Beta-amyloid protein increases the vulnerability of cultured cortical neurons to excitotoxic damage, Brain Research, (1990), 533, p315-320.
- Kontush, A, Amyloid-beta: an antioxidant that becomes a pro-oxidant and critically contributes to Alzheimer's disease, Free radical Biology & Medicine, (2001), 31, p1120-1131.
- Kumar, S, Sharma, A, Madan, B, Singhal, V, Ghosh, B, Isoliquiritigenin inhibits IkappaB kinase activity and ROS generation to block TNF-alpha induced expression of cell adhesion molecules on human endothelial cells, Biochemical Pharmacology, (2007), 73, p1602-1612.
- Lee, BY, Ban, JY, Seong, YH, Chronic stimulation of GABAA receptor with muscimol reduces amyloid beta protein(25-35)-induced neurotoxicity in cultured rat cortical cells, Neuroscience Research, (2005), 52, p347-356.
- Lee, HK, Yang, EJ, Kim, JY, Song, KS, Seong, YH, Inhibitory effects ofGlycyrrhizaeradix and its active component isoliquiritigenin on Abeta(25-35)-induced neurotoxicity in cultured rat cortical neurons, Archives of Pharmacal Research, (2012), 35, p897-904.
-
Lee, IS, Jin, W, Zhang, X, Hung, TM, Song, KS, Seong, YH, Bae, K, Cytotoxic and COX-2 inhibitory constituents from the aerial parts ofAralia cordata, Archives of Pharmacal Research, (2006), 29, p548-555.
[https://doi.org/10.1007/bf02969263]
- Maurice, T, Lockhart, BP, Privat, A, Amnesia induced in mice by centrally administered beta-amyloid peptides involves cholinergic dysfunction, Brain Research, (1996), 706, p181-193.
- Miranda, S, Opazo, C, Larrondo, LF, Munoz, FJ, Ruiz, F, Leighton, F, Inestrosa, NC, The role of oxidative stress in the toxicity induced by amyloid beta-peptide in Alzheimer's disease, Progress in Neurobiology, (2000), 62, p633-648.
- Morais, Cardoso S, Swerdlow, RH, Oliveira, CR, Induction of cytochrome c-mediated apoptosis by amyloid beta 25-35 requires functional mitochondria, Brain Research, (2002), 931, p117-125.
-
Park, HJ, Hong, MS, Lee, JS, Leem, KH, Kim, CJ, Kim, JW, Lim, S, Effects ofAralia continentalison hyperalgesia with peripheral inflammation, Phytotherapy Research, (2005), 19, p511-513.
[https://doi.org/10.1002/ptr.1693]
- Pike, CJ, Walencewicz-Wasserman, AJ, Kosmoski, J, Cribbs, DH, Glabe, CG, Cotman, CW, Structure-activity analyses of beta-amyloid peptides: Contributions of the beta 25-35 region to aggregation and neurotoxicity, Journal of Neurochemistry, (1995), 64, p253-265.
-
Rice-Evans, CA, Miller, NJ, Bolwell, PG, Bramley, PM, Pridham, JB, The relative antioxidant activities of plant-derived polyphenolic flavonoids, Free Radical Research, (1995), 22, p375-383.
[https://doi.org/10.3109/10715769509145649]
- Sano, M, Ernesto, C, Thomas, RG, Klauber, MR, Schafer, K, Grundman, M, Woodbury, P, Growdon, J, Cotman, CW, Pfeiffer, E, Schneider, LS, Thal, LJ, A controlled trial of selegiline alpha-tocopherol or both as treatment for Alzheimer's disease. The Alzheimer's Disease Cooperative Study, The New England Journal of Medicine, (1997), 336, p1216-1222.
-
Smith, MA, Richey Harris, PL, Sayre, LM, Beckman, JS, Perry, G, Widespread peroxynitrite-mediated damage in Alzheimer's disease, The Journal of Neuroscience, (1997), 17, p2653-2657.
[https://doi.org/10.1523/jneurosci.17-08-02653.1997]
- Stepanichev, MY, Moiseeva, YV, Lazareva, NA, Onufriev, MV, Gulyaeva, NV, Single intracerebroventricular administra n of amyloid-beta(25-35) peptide induces impairment in short-term rather than long-term memory in rats, Brain Research Bulletin, (2003), 61, p197-205.
-
Tang, SY, Whiteman, M, Peng, ZF, Jenner, A, Yong, EL, Halliwell, B, Characterization of antioxidant and antiglycation properties and isolation of active ingredients from traditional chinese medicines, Free radical Biology & Medicine, (2004), 36, p1575-1587.
[https://doi.org/10.1016/j.freeradbiomed.2004.03.017]
- Tohda, C, Matsumoto, N, Zou, K, Meselhy, MR, Komatsu, K, Abeta(25-35)-induced memory impairment axonal atrophy and synaptic loss are ameliorated by M1 A metabolite of protopanaxadiol-type saponins, Neuropsychopharmacology, (2004), 29, p860-868.
- Ueda, K, Shinohara, S, Yagami, T, Asakura, K, Kawasaki, K, Amyloid beta protein potentiates Ca2+influx through Ltype voltage-sensitive Ca2+channels: A possible involvement of free radicals, Journal of Neurochemistry, (1997), 68, p265-271.
- Um, MY, Choi, WH, Aan, JY, Kim, SR, Ha, TY, Protective effect ofPolygonum multiflorumThunb on amyloid beta-peptide 25-35 induced cognitive deficits in mice, Journal of Ethnopharmacology, (2006), 104, p144-148.
-
Wang, Q, Yu, S, Simonyi, A, Rottinghaus, G, Sun, GY, Sun, AY, Resveratrol protects against neurotoxicity induced by kainic acid, Neurochemical Research, (2004), 29, p2105-2112.
[https://doi.org/10.1007/s11064-004-6883-z]
- Yan, XZ, Qiao, JT, Dou, Y, Qiao, ZD, Beta-amyloid peptide fragment 31-35 induces apoptosis in cultured cortical neurons, Neuroscience, (1999), 92, p177-184.